


Abstrakt
Centrálna dogma genetiky hovorí, že prenos informácie v živej bunke je možný len jediným smerom: z DNA cez RNA do proteínu. Objav enzýmov reverzných transkriptáz však dokázal, že z RNA je možný spätný zápis do DNA. Ja som sa pokúsila túto spätnú cestu prenosu informácie skompletizovať a to návrhom teoretického riešenia prenosu informácie z proteínu do RNA, čiže reverznej translácie (retrotranslácie). Reverzná translácia by sa uskutočňovala v špecifických bunkových organelách nazývaných retrozómy (skrátenie názvu retroribozómy). Retrozóm by zastával opačnú funkciu ako ribozóm: kým v ribozóme dochádza k čítaniu molekuly mRNA a na základe informácie v nej obsiahnutej k zostaveniu proteínového reťazca z monomérov – aminokyselín, v retrozóme by bol proteínový reťazec rozložený na jednotlivé aminokyseliny a na základe nich by sa zostavila molekula RNA. Túto molekulu RNA, výsledok procesu retrotranslácie, budeme nazývať cRNA. cRNA je čo do kódujúcej oblasti totožná s mRNA príslušného génu, ale obsahovala by navyše matrice pre regulačné sekvencie, ktoré sa z pre-mRNA odstraňujú zostrihom. Ďalej by obsahovala sekvencie, ktoré by umožnili transport cRNA do jadra a prepis do formy normálneho génu na vhodné miesto genómu. Retrotranslácia je hypotéza nepodložená akýmkoľvek pozorovaním; zostavila som ju len ako myšlienkový experiment, špekuláciu, ktorá, pokiaľ jej fyzikálne a chemické zákony nebránia, mohla evolučne vzniknúť, ale nevznikla.
Úvod
V každej živej sústave je možné zlepšovanie efektívnosti životných pochodov a preto každá živá sústava podlieha pomalému vývoju – evolúcii. Základom evolúcie sú postupné zmeny v DNA, ktoré sú o to pomalšie, že väčšina náhodných zmien v DNA je opravovaná bunkou samotnou. Vývoj úplne nového génu je preto nesmierne pomalý proces, ktorý zahŕňa tisíce, stotisíce až milióny náhodných mutácií na určitých miestach. Tieto mutácie musia vytvoriť nielen kód pre zapojenie aminokyselín do funkčného celku, ale aj regulačné sekvencie pre expresiu tohto génu. Z tohto pohľadu je retrotranslácia úžasnou evolučnou skratkou. Jej výsledkom by bol totiž vznik plne funkčného génu so správnym poradím báz vytvárajúcim kodóny, so všetkými regulačnými sekvenciami, v priebehu niekoľkých hodín. Samozrejme, takýto gén by potom tiež podliehal evolúcii a postupnému pomalému zlepšovaniu.
Spúšťačom retrotranslácie by bola prítomnosť nového proteínu v bunke, proteínu, pre ktorý bunka nemá (funkčný) gén, ale ktorý plní svoju funkciu lepšie ako vlastné proteíny bunky, prípadne má celkom novú, ale osožnú funkciu. Spôsob rozpoznania takéhoto proteínu bunkou a zhodnotenie jeho užitočnosti pre bunku už leží mimo tejto úvahy – nepochybne by to vyžadovalo zložitú signálnu dráhu navyše komplikovanú tým, že bunka sa proteínov, ktoré nie sú jej vlastné, snaží v prvom rade zbaviť. Moja úvaha začína v bode, kedy bunka cudzí proteín už vyhodnotila ako výhodný pre ňu a rozhodla sa vytvoriť si jeho gén – čiže nastalo spustenie procesu retrotranslácie, po ktorom nasleduje proces retrotranskripcie.
Na retrotransláciu sú potrebné organely retrozómy. Výsledkom retrotranslácie je kodónová RNA (cRNA) pripravená na reverznú transkripciu do DNA. Proces retrotranslácie si vyžaduje dva druhy RNA, ktoré budeme nazývať kyselina retrotransferová (rtRNA) – tá je analógiou tRNA, a kyselinu prekodónovú (pcRNA), ktorá je predchodcom budúcej cRNA. Tieto dva druhy molekúl by sa pohybovali voľne v cytoplazme. Nemožno opomenúť ďalšie štyri hlavné skupiny enzýmov retrotranslácie: exoproteáza schopná odštepovať vždy len koncovú aminokyselinu z C- konca peptidu, aminoacyl tRNA syntetázy, RNA-ligáza, a restrikčná endonukleáza, ktorej celá rozpoznávacia sekvencia leží na 5´ konci od štiepiaceho miesta. Všetky tieto komponenty by boli kódované genómom bunky.
Komponenty
Retrozóm
Retrotranslácia by prebiehala v bunkových organelách zvaných retrozómy, ktorých by, vzhľadom na zriedkavú potrebu, bunka obsahovala omnoho menej ako ribozómov. Retrozóm by bol, podobne ako ribozóm, komplex zložený z proteínov a RNA, ktorý sa však od ribozómu odlišuje nevyhnutným napojením na membránovú organelu. Pri malom rozlíšení v elektrónovom mikroskope by retrozómy mohli pripomínať ribozómy prisadnuté na membráne endoplazmatického retikula. Membránou uzavretý priestor potrebný pre retrozóm by však bol podstatne menší, ako v prípade pravého ER a mohol by sa približovať k limitnej hodnote, akú membránová organela vzhľadom na usporiadanie svojich stavebných zložiek môže mať, nakoľko na retrotransláciu je potrebné len veľmi malé množstvo cieľového proteínu (stačí jedna molekula s kompletným neporušeným pôvodným aminokyselinovým reťazcom). Nukleoproteínový komplex retrozómu by obsahoval dutinu, v ktorej by prebiehal vlastný proces retrotranslácie. Dutina by bola spojená s cytoplazmou. Nepostrádateľnou súčasťou retrozómu je tiež základ cRNA. Tým základom je RNA, ktorá nie je nedeliteľnou súčasťou nukleoproteínového komplexu retrozómu a ktorej 5´ koniec je zasunutý v dutine retrozómu, kým 3´ koniec prečnieva voľne von mimo retrozomálneho komplexu. Táto RNA je budúci 3´ koniec cRNA.
Retrotransferová ribonukleová kyselina
Podobne ako v procese translácie, aj pri retrotranslácii má kľúčovú úlohu adaptorová molekula RNA, ktorá je sprostredkovateľom informácie medzi aminokyselinou a príslušným kodónom. RNA, ktorá má túto úlohu v procese translácie, budem nazývať retrotransferová ribonukleová kyselina, skrátene rtRNA. Počet verzií rtRNA v bunke je rovnaký ako počet verzií tRNA a 3´ konce každého druhu rtRNA majú svoj identický náprotivok v 3´ koncoch tRNA. 5´ koniec rtRNA sa však od 5´ konca tRNA značne líši. 5´ konce rtRNA nie sú otočené naspäť a nepárujú sa s nijakou sekvenciou v rámci jednej rtRNA ani medzi rôznymi rtRNA navzájom. Ide o rozdielne, pre každú rtRNA špecifické sekvencie, ktoré obsahujú výlučne alebo takmer výlučne guanín a cytozín.
Prekodónová ribonukleová kyselina
Druhou, nemenej významnou RNA v procese retrotranslácie je takzvaná prekodónová RNA, v skratke pcRNA. Ak máme hľadať jej analóg v procese translácie, tak je ním samotná mRNA. pcRNA totiž už nesú kodóny, ktoré sa stanú súčasťou RNA, každá pcRNA jeden a sú to posledné tri nukleotidy jej 3´ konca. Za touto trojicou nukleotidou smerom k 5´ koncu sa nachádza sekvencia, ktorú majú všetku druhy pcRNA identickú, pretože slúži ako rozpoznávacie miesto pre restrikčnú endonukleázu. Za touto identickou sekvenciou leží oblasť bohatá na guanín a cytozín, ktorou sa každá jedna pcRNA presne a pevne páruje s príslušnou rtRNA, ktorá k nej sekvenčne pasuje a ktorej 3´ koniec viažuci aminokyselinu zodpovedá správnemu kodónu na 3´ konci pcRNA. Každá pcRNA má teda tri dôležité úseky: smerom od 3´ konca sú to kodón, identická sekvencia pre restriktázu a oblasť guanínu a cytozínu slúžiaca na naviazanie sa na príslušnú rtRNA. Sekvencie pcRNA sú zároveň stavané tak, že molekule neumožňujú zaujať nijakú stabilnú sekundárnu štruktúru – v cytoplazme sa teda pohybuje „vystretá“.
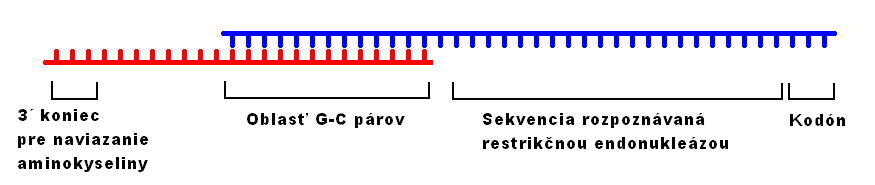
Obrázok 1: topológia rtRNA (červená) a pcRNA (modrá). Počet tu znázornených báz nezodpovedá skutočnosti.
Prípravná fáza
Proteín určený na retrotransláciu musí byť zložený iba z jedného peptidového reťazca – viacero reťazcov môže byť začlenených do genómu viacerými procesmi retrotranslácie. Na začiatku procesu je cieľový proteín transportovaný do membránovej časti retrozómu. V tomto prostredí je postupne zbavený všetkých naviazaných komponentov (kovov a i.) a je tiež odstránená jeho terciálna aj sekundárna štruktúra, pričom ale nesmie byť narušená primárna štruktúra. Tento proces môže prebiehať postupne aj v niektorých navzájom oddelených membránových váčkoch, medzi ktorými sú čiastkové produkty premiestňované vo vezikulách podobne ako v Golgiho aparáte. Výsledkom celého procesu je odstrojený neposkladaný lineárny peptidový reťazec. Pokiaľ ho už jeho primárna štruktúra predurčuje k určitej sekundárnej štruktúre neodstrániteľnej ani iným chemickým zložením a pH vo vnútri membránovej organely, na peptide musia byť naviazané určité proteíny, ktoré bránia vytvoreniu väzieb potrebných napríklad pre štruktúru alfa helixu. Tieto proteíny však nemusia byť umiestnené súčasne po celej dĺžke peptidového reťazca – stačí, aby „vyrovnaný“ zostal jeho C- koniec a s postupujúcou retrotransláciou sa budú vyrovnávacie proteíny posúvať ďalej smerom k N- koncu.
Do retrotranslácie vstupuje vždy najdlhší peptidový reťazec cieľového proteínu, pretože ten sa považuje za najmenej (respektíve vôbec) degradovaný. Ostatné, kratšie reťazce s rovnakými úsekmi aminokyselinových sekvencií sú nepotrebné, po ukončení retrotranslačného procesu sú degradované a ich stavebné komponenty sú transportované preč z membránového priestoru retrozómu.
Priebeh retrotranslácie
Voľný C- koniec cieľového peptidu sa vsúva do tunela nukleoproteínovej časti retrozómu podobne, ako sa po translácii koniec vznikajúceho proteínu vysúval z ribozómu. Proces retrotranslácie je však otočeným procesom translácie a namiesto vzniku dôjde postupne k úplnej degradácii peptidového reťazca za súčasného vzniku cRNA. Na základe jedného peptidového reťazca je preto možné utvoriť len jednu cRNA, čím sa tento proces líši od translácie, lebo pri translácii možno podľa jednej mRNA vytvoriť niekoľko peptidových reťazcov. Systém jeden peptid – jedna cRNA je však výhodný z toho hľadiska, aby nedochádzalo k tvorbe nadbytočných kópií daného génu.
C koniec peptidu je teda vsunutý do tunelu retrozómu. Tam na neho čaká proteáza kovalentne naviazaná k retrozómu a odštepujúca výhradne koncové aminokyseliny. Táto proteáza odštiepi z C- konca prvú aminokyselinu. Voľná aminokyselina sa pohybuje v malej dutine, do ktorej z cytoplazmy prúdia komplexy zložené z dvoch RNA: rtRNA s pcRNA a enzýmov aminoacyl tRNA-syntetáz.
Príslušná aminoacyl tRNA syntetáza rozpozná trojbázovú sekvenciu na 3´ konci správnej rtRNA a pripojí k tomuto koncu aminokyselinu. rtRNA slúži ako adaptor a nosič pcRNA. Najdôležitejšou časťou pcRNA sú tri bázy na jej 3´ konci – kodón. Keď dôjde k naviazaniu aminokyseliny na 3´ koniec rtRNA, celý komplex aminokyselina-rtRNA-pcRNA prejde konformačnými zmenami, ktoré umožnia pcRNA spojiť sa kodónom s 5´ koncom základu cRNA.
RNA základ cRNA je v retrozóme už od iniciácie retrotranslácie. Obsahuje sekvencie, ktoré po prepise do DNA budú slúžiť ako regulačné sekvencie pre transkripciu. Keďže cRNA sa bude skladať od 3´ konca smerom k 5´ koncu, začiatočný úsek cRNA obsahuje sekvencie, ktoré po prepise reverznou transkriptázou budú zodpovedať terminátoru. Na tento základ sa naviaže voľný, čiže 3´ koniec pcRNA. Molekuly spojí RNA-ligáza. Tak sme dostali na správne miesto prvú trojicu báz tvoriacu posledný kodón budúceho génu. Avšak za ním sa stále nachádza teraz už neužitočná sekvencia pcRNA, navyše stále spárovaná s rtRNA. Tento problém vyrieši restrikčná endonukleáza, ktorá rozštiepi pcRNA hneď za kodónom vďaka sekvencii, ktorá jej dáva na štiepenie signál. Kodón je teda úspešne pripojený, zvyšok komplexu pcRNA-rtRNA-aminokyselina opúšťa dutinu retrozómu.
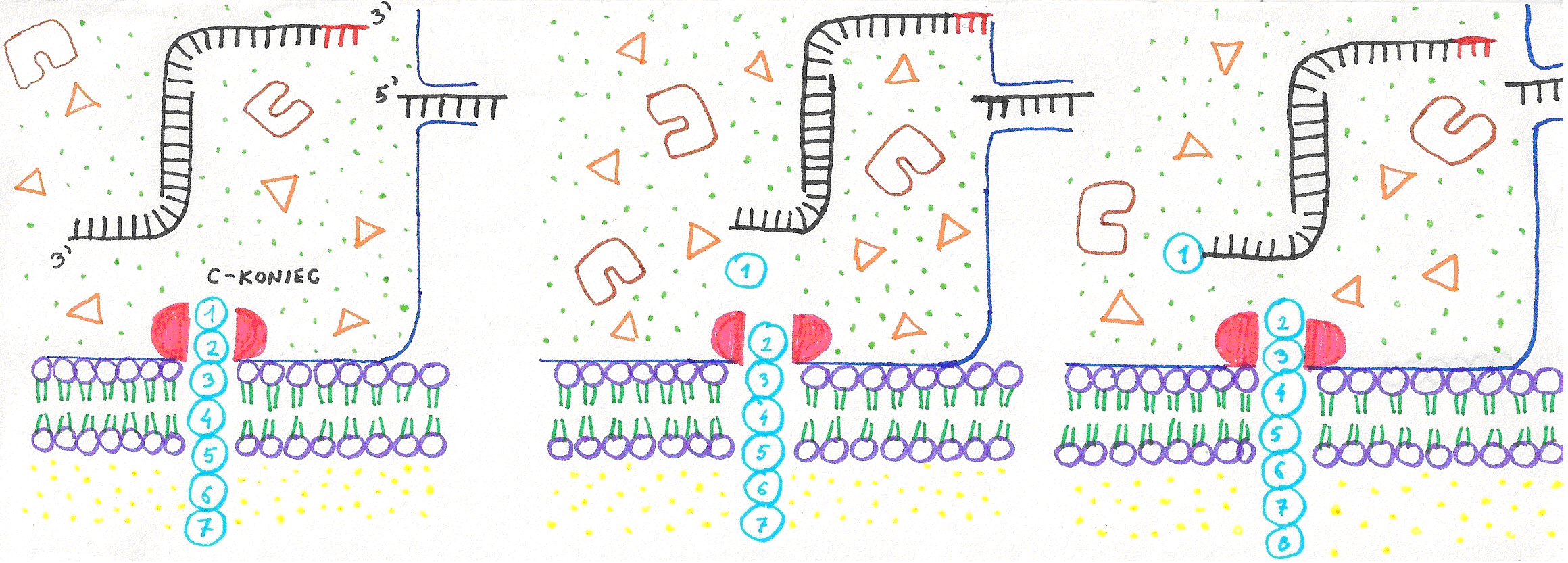
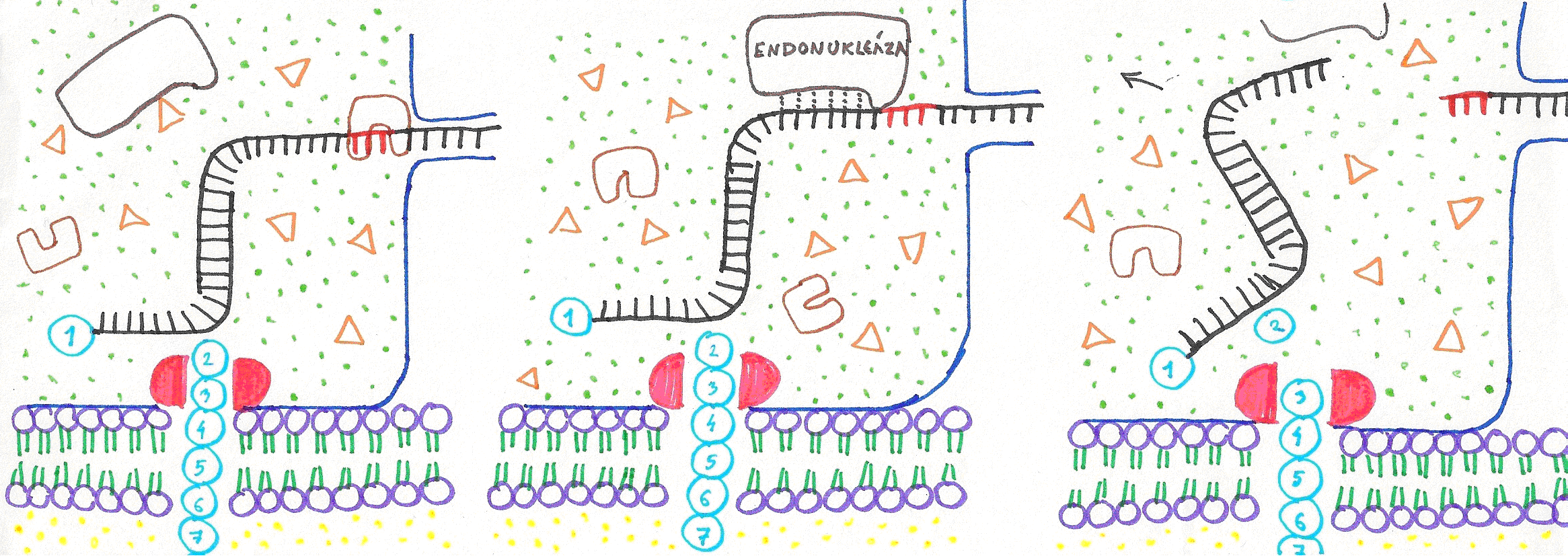
Obrázok 2: Priebeh retrotranslácie:
- C-koniec proteínu sa vsunie do nukleoproteínovej časti retrozómu
- proteáza odštiepi C-koncovú aminokyselinu
- Aminoacyl tRNA sysntetáza pripojí aminokyselinu k 3´ koncu rtRNA
- pcRNA sa kodónom pripojí k základu cRNA
- restrikčná endonukleáza odštiepi zvyšok pcRNA za kodónom
- rtRNA so zvyškom pcRNA opúšťajú dutinu retrozómu. Proteáza odštepuje druhú C-koncovú aminokyselinu.
Z C- konca peptidu je odštiepená ďalšia aminokyselina. Tá si počká na príslušnú rtRNA, na ktorej 3´ koniec sa naviaže a celý proces sa opakuje, až kým nie je peptidový reťazec úplne degradovaný a všetky kodóny skompletizované. Následne sa k novovznikajúcej cRNA pripojí RNA obsahujúca regulačné sekvencie na jej 5´ konci, čiže negatív promótora. Takáto RNA je pripravená na transport do jadra, kde bude reverznou transkriptázou prepísaná do formy DNA. Samozrejme, gén vzniknutý týmto spôsobom sa bude stále v mnohom odlišovať od génov vzniknutých pomalou evolúciou – nebude obsahovať žiadne intróny. Ani táto odlišnosť však v princípe nie je neprekonateľná. Pred reverznou transkripciou môže cRNA v jadre podstúpiť ešte „predtranskripčné modifikácie“, medzi ktoré by patril aj insert – opak splicingu. V priebehu insertu by hypotetický enzým intráza s vlastnosťami rekombinázy vsúval do kódujúcej oblasti intróny. Vložené intróny by boli RNA úseky kódované genómom bunky so sekvenciami zabezpečujúcimi budúcemu génu úspešný splicing.
Problémy
Problém štart a stop kodónu
Nesmieme zabúdať na fakt, že okrem regulačných sekvencií je pre úspešnú transláciu kľúčový aj prvý kodón na 5´ konci mRNA, tzv. štart kodón. Vieme, že štart kodón zodpovedá aminokyseline metionínu. Problém je však v tom, že v mnohých prípadoch bol metionín v priebehu posttranslačných modifikácií odštiepený. Metionín bude preto chýbať, pri retrotranslácii sa nebude môcť naviazať na príslušnú rtRNA a nebude môcť pridať posledný kodón – iniciačný. To síce retrotransláciu nijako neovplyvní, ale omnoho neskôr, po vytvorení transkriptu z génu vzniknutého vďaka retrotranslácii nebude možné takýto gén translatovať – celý proces retrotranslácie by bol teda zbytočný.
Tento problém môže byť vyriešený tak, že v už predpripravenej sekvencii tvoriacej budúci 5´ koniec cRNA sa bude iniciačný kodón nachádzať. Budúca translácia preto už nebude závislá od prítomnosti metionínu na N- konci peptidového reťazca. Pokiaľ sa náhodou metionín na N- konci vzorového peptidového reťazca ešte nachádzal aj v čase retrotranslácie, nie je to prekážkou – v reťazci cRNA tak budú dva iniciačné kodóny (AUG) za sebou, čo translácii v budúcnosti nezabráni. Pri posttranslačných modifikáciách peptidu však bude potrebné nadbytočný metionín odštiepiť.
Podobným spôsobom by sa dal vyriešiť aj problém chýbajúceho stop kodónu, ktorý, keďže mu nepripadá nijaká aminokyselina, nemôže byť prinesený na vznikajúci cRNA reťazec prostredníctvom adaptorovej molekuly rtRNA. Základ cRNA, 3´ koniec, teda v sebe už stop kodón musí obsahovať.
Problém degenerácie genetického kódu
Kým pri translácii ku každej matrici-kodónu môže byť priradená iba jedna aminokyselina, pri retrotranslácii je kvôli degenerácii genetického kódu možné priradiť k niektorým aminokyselinám niekoľko kodónov. To sa dá vyriešiť tým, že pcRNA pri retrotranslácii môžu obsahovať náhodné synonymné kodóny, alebo budú preferovať či obsahovať výlučne len jeden kodón, ktorý je daným organizmom najpoužívanejší.

Napísať odpoveď