


Prvýkrát publikované na slovenskej Wikipédii. Autor: Jana Plauchová ako Eryn Blaireová. Spoluautori: FILIT – zdroj, z ktorého pôvodne čerpal základ článku, Vojtěch Dostál (časť textu, česká Wikipédia), IP 89.102.219.85 (časť textu, česká Wikipédia), IP 84.245.71.66 (oprava gramatiky), Abyss (doplnenie informácií), Improbus (úprava formulácie a oprava preklepov), IP 193.87.105.213 (oprava preklepu)
Text je dostupný pod Creative Commons Attribution-ShareAlike License 3.0 a GFDL.

Časť ribozómu; bielkoviny sú znázornené namodro
Ribozómy sú bunkové organely nachádzajúce sa vo všetkých známych bunkách, čiže vo všetkých živých organizmoch okrem vírusov, pokiaľ sú pokladané za živé. Prebieha v nich syntéza bielkovín (proteínov) – proteosyntéza. Patria medzi najhojnejšie zastúpené bunkové organely. Sú uložené voľne v cytoplazme, u eukaryotov sú tiež vo vnútri plastidov, mitochondrií a viazané na endoplazmatické retikulum. Obsahujú ribonukleovú kyselinu (RNA) a bielkoviny, z chemického hľadiska preto patria medzi nukleoproteíny.
Bielkoviny tvoria cca 1/3 obsahu ribozómu. Odlišujeme dva základné typy týchto organel (pre prokaryoty a eukaryoty). Eukaryoty však majú v semiautonómnych organelách ribozómy podobné prokaryotickým ribozómom a iba ich cytoplazmatické ribozómy sa označujú ako ribozómy eukaryotického typu.
Každý ribozóm sa delí na dve podjednotky s odlišnými veľkosťami. K ribozómu sa napája tzv. mediátorová ribonukleová kyselina (mRNA), ktorá obsahuje prepis genetickej informácie z deoxyribonukleovej kyseliny (DNA). Ribozóm potom podľa tejto informácie nasyntetizuje polypeptid resp. proteín. Proces jeho syntézy sa nazýva translácia a prebieha tak, že ribozóm sa pohybuje pozdĺž mRNA a prekladá jej kodóny do peptidového reťazca. Po pripojení každej aminokyseliny k peptidovému reťazcu dochádza k posunu ribozómu po mRNA o tri nukleotidy (kodón) smerom k 3´ koncu, čo sa označuje ako translokácia ribozómu. V prokaryotických bunkách sa ribozóm pohybuje po mRNA rýchlosťou 30 – 60 nukleotidov za sekundu, v eukaryotických je to len 9 až 15 nukleotidov/sek.
Počet ribozómov v bunkách je veľmi variabilný. V baktérii Escherichia coli ich napríklad je priemerne asi 15 000. Jedna molekula mRNA môže byť súčasne translatovaná viacerými ribozómami. V prípade, že je na jednu mRNA nadviazaných viac ribozómov, hovoríme o polyribozóme alebo o polyzóme.
Vznik
Ribozómy sú zložené z ribozomálnych proteínov a z ribozomálnej RNA (rRNA). Ribozomálne proteíny sa syntetizujú na už existujúcich ribozómoch v cytoplazme bunky. Po ich vytvorení sú tieto proteíny v eukaryotických bunkách dopravené do jadra bunky. Tam sa spájajú s novosyntetizovanou rRNA už pred dokončením jej vzniku transkripciou. Takto sa skopmletizujú celé podjednotky ribozómov a následne sú dopravené opäť do cytoplazmy, kde sa zúčastňujú proteosyntézy.
Gény pre rRNA sú najaktívnejšie prepisované vtedy, keď (bakteriálna) bunka podstupuje rýchle delenie, ktoré je prirodzene spojené s veľkou potrebou proteínov. V rýchlo rastúcich bakteriálnych kultúrach môžu tvoriť ribozómy aj viac ako tretinu ich hmoty. Transkripciu rRNA u baktérií reguluje guanozín-5´-bisfosfát-3´bisfosfát, skrátene ppGpp. Pri nedostatku ppGpp intenzívne prebieha syntéza rRNA, ale pri jeho nadbytku sa zastaví. ppGpp pravdepodobne pôsobí na RNA-polymerázu, ktorá v jeho prítomnosti stráca afinitu k promótoru génu pre rRNA.
Stavba ribozómu
Ribozóm je malá bunková organela s priemerom 17 až 20 nm, preto je pozorovateľný len elektrónovým mikroskopom. Možno ich z bunky separovať mechanickým rozdrvením bunky skleným práškom alebo enzymatickým rozrušením bunkovej steny. V rastlinných a živočíšnych bunkách sú všetky ribozómy prisadnuté na membránach.
Ribozóm sa približne z dvoch tretín skladá z ribonukleovej kyseliny (konkrétne tzv. rRNA, teda ribozomálnej RNA), a zvyšnú tretinu tvoria proteíny. Všetky organizmy majú podobnú stavbu ribozómov. Bielkoviny a molekuly rRNA sú navzájom spojené iónovými väzbami a vodíkovými väzbami. U prokaryotov sa ribozómy skladajú z 65 % rRNA a 35 % proteínov, ribozómy endoplazmatického retikula eukaryotov obsahujú 55 % rRNA a 45 % proteínov.
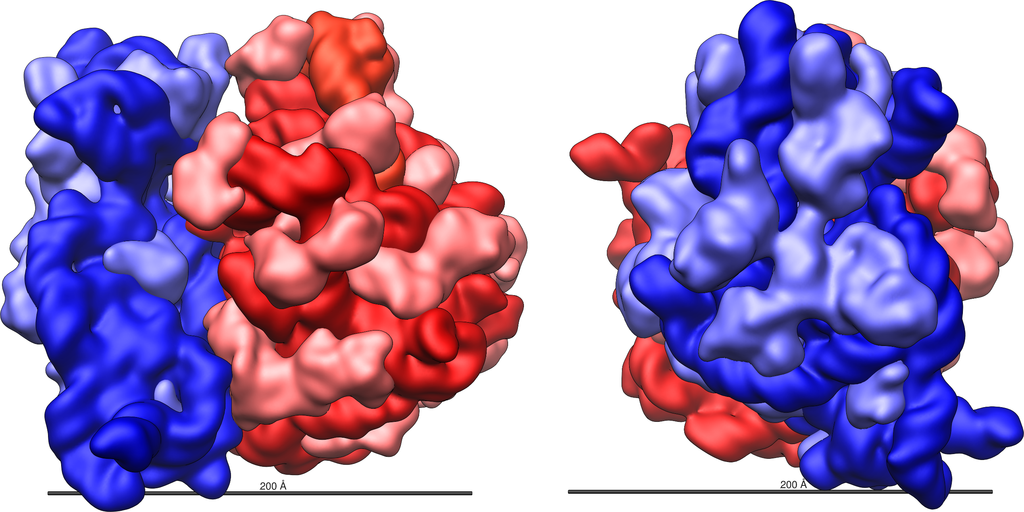
Veľká (červená) a malá (modrá) podjednotka ribozómu baktérie Escherichia coli
Všetky ribozómy možno rozdeliť na dve základné časti, na malú a veľkú podjednotku (subjednotku). Napriek tomu však existujú určité rozdiely, hlavne v stavbe, medzi prokaryotickými a eukaryotickými ribozómami. Pod pojmom eukaryotický ribozóm sa myslia ribozómy umiestnené mimo plastidov a mitochondrií. Ribozómy plastidov a mitochondrií sa totiž podobajú prokaryotickým ribozómom, z ktorých pravdepodobne vznikli, svojou sedimentačnou konštantou aj spôsobom translácie.
Rozdiely medzi ribozómami sa veľmi často udávajú pomocou tzv. sedimentačného koeficientu, čiže veličiny, ktorá udáva čas, za ktorý v ultracentrifúge prebehne sedimentácia ribozómu. Jednotkou je Svedberg (S), a táto jednotka predstavuje čas 10−13 sekundy. Prokaryotický ribozóm sa na základe týchto veličín označuje ako 70S, eukaryotický je 80S. Tiež obe podjednotky vykazujú určité rozdiely, pokiaľ porovnáme sedimentačné koeficienty eukaryotických a prokaryotických ribozómov. Taktiež ribozomálna RNA sa rozlišuje na základe sedimentácie; u prokaryotov poznáme 5S, 16S a 23S rRNA.
Podjednotky sú spolu a utvárajú ribozóm len počas proteosyntézy. Po jej skončení sa rozpadajú na samostatné podjednotky, ktoré sú zrejme až do ďalšieho spojenia s mRNA v bunke vo voľnom stave. Na podjednotky ribozómy disociujú napríklad za zníženej koncentrácie iónov draslíka. Spojeniu oboch jednotiek u prokaryotov bráni iniciačný faktor IF3, ktorý sa naviaže na malú podjednotku. Tento faktor ale zároveň umožňuje, aby sa naň nadviazal komplex zložený z iniciačného faktoru IF2, guanozíntrifosfátu (GTP) a tRNA nesúcej formylovaný metionín. Malá podjednotka je teda pripravená zahájiť iniciáciu translácie.
Malá podjednotka
Úlohou malej podjednotky je nasadiť tRNA na kodóny mRNA v procese translácie. Malá podjednotka prokaryotov má koeficient 30S, u eukaryotov je táto podjednotka 40S. Za začiatok translácie peptidového reťazca sa považuje okamih, keď sa táto malá podjednotka naviaže na mRNA. To sa deje skôr ako nadviazanie veľkej podjednotky. Pripojenie veľkej podjednotky je umožnené len po nadviazaní mRNA a prvej (iniciačnej) molekuly transferovej ribonukleovej kyseliny (tRNA). Tieto udalosti spôsobia zmenu konformácie malej podjednotky. Malá podjednotka v priebehu translácie sprostredkováva dekódujúcu interakciu medzi kodónom na mRNA a antikodónom na tRNA.
Veľká podjednotka
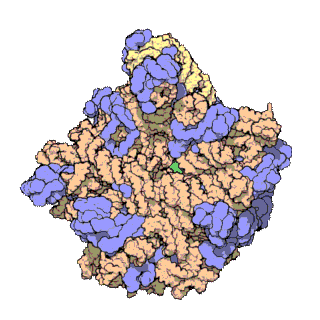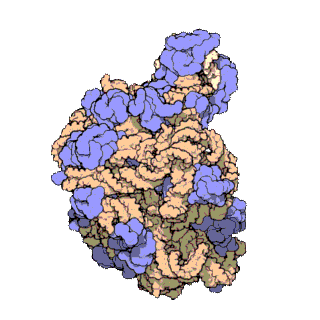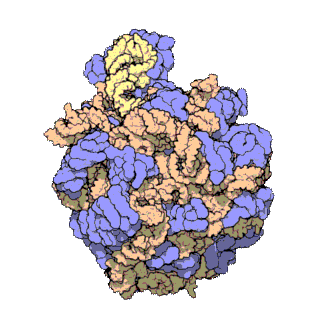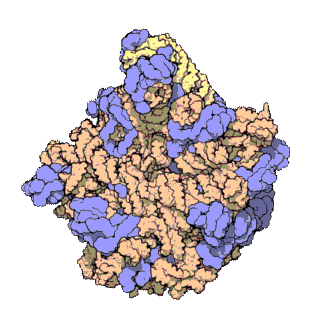
Veľká ribozomálna podjednotka archebaktérie Haloarcula marismortui
Veľká podjednotka slúži počas translácie na katalyzovanie vzniku peptidovej väzby medzi aminokyselinou a vznikajúcim reťazcom. Veľká podjednotka ribozómu je u prokaryotov 50S, u eukaryotov 60S. Veľká podjednotka sa pripája na tzv. prediniciačný komplex zložený z iniciačných faktorov, RNA a malej podjednotky. Spojením veľkej a malej podjednotky sa ribozóm kompletizuje a vzniká tzv. iniciačný komplex, ktorý prechádza v translácii do fázy elongácie. Podjednotky sa spájajú v blízkosti 5´ konca mRNA. Vo veľkej podjednotke je akýsi „tunel“, v ktorom zostáva skrytý rastúci peptidový reťazec, až kým nedosiahne dĺžku približne 70 aminokyselín. Až vtedy sa jeho N-koniec vynára von z ribozómu.
Väzobné miesta
V ribozóme sa nachádzajú celkove 4 väzobné miesta pre molekuly RNA: jedno pre mRNA a tri pre aktivované formy tRNA (aminoacyl-tRNA, skrátene aa-tRNA). Väzobné miesto pre mRNA sa nachádza na malej podjednotke, komplexy aminoacyl-tRNA sa z veľkej časti viažu na styčnom mieste oboch podjednotiek. Táto styčná plocha je bohatšia na rRNA a chudobnejšia na ribozómové bielkoviny ako ostatné časti ribozómu. Miesta pre tRNA sú označené ako aminoacylové (A), peptidylové (P) a výstupné (E) miesto. Aminoacylové miesto sa nachádza z menšej časti na malej podjednotke, z väčšej časti na väčšej podjednotke a vstupujú doň aa-tRNA. Odtiaľ sa presunú na pepitdylové miesto, ktoré je, podobne ako A miesto, lokalizované z menšej časti na menšej podjednotke a z väčšej časti na väčšej podjednotke. Na toto miesto sa viaže tRNA, ktorej aminokyselina je už spojená so vznikajúcim peptidovým reťazcom, ale stále pretrváva aj esterová väzba medzi 3´ tRNA, ktorá najnovšiu aminokyselinu doniesla. Po oddelení sa tRNA od aminokyseliny tRNA postupuje na E miesto, ktorého názov je odvodený z anglického slova exit (východ), pretože cezeň deacetylovaná tRNA opúšťa ribozóm. Vznik peptidovej väzby medzi prichádzajúcimi aminokyselinami sa uskutočňuje na peptidyltransferázovom mieste.
Ďalej sa na ribozóme nachádzajú väzobné miesta pre ďalšie nevyhnutné súčasti translácie – iniciačné a elongačné faktory. Vnútorná hmota ribozómu sa označuje ako dreň (core).
rRNA
Ribozomálna RNA je esenciálnou zložkou ribozómov. Zistilo sa, že práve rRNA je zodpovedná za funkčnosť ribozómu, čiže jeho schopnosť prepisovať mRNA do proteínov. Z tohto pohľadu je ribozomálna RNA vlastne enzým a preto sa nazýva ribozým. Objav tejto funkcie bol spočiatku prekvapivý, pretože sa myslelo, že enzymatickej aktivity sú schopné len proteíny. Ribozomálna RNA však tvorí priestorové štruktúry, podobné aktívnym miestam proteínov fungujúcich ako enzýmy. Vďaka tomu je schopná napríklad správne nadviazať aa-tRNA a taktiež zaisťuje vznik peptidových väzieb medzi aminokyselinami vznikajúceho reťazca (druhú z menovaných ovláda u prokaryotov predovšetkým 23S rRNA).
Zjednodušená schéma priebehu translácie v ribozóme: mRNA (čierna lineárna štruktúra) sa stretáva s malou podjednotkou (žltá štruktúra). Následne sa pridáva veľká podjednotka (zelená štruktúra) a aminoacyltRNA, ktoré prinášajú aminokyseliny pre postupne budovaný proteínový reťazec. Na ribozóme prisadnutom na membráne endoplazmatického retikula (pokračovanie animácie) sa novovytváraný peptidový reťazec zanára rovno dovnútra endoplazmatického retikula, aby v ňom podstúpil ďalšie úpravy.
Prokaryotický a eukaryotický ribozóm sa však v obsahu rRNA líšia. Zatiaľ čo prokaryotický obsahuje v malej podjednotke 16S rRNA (1540 nukleotidov) a vo veľkej podjednotke 5S rRNA (120 nukleotidov) a 23S rRNA (2900 nukleotidov), eukaryotický obsahuje v malej podjednotke 18S rRNA (1 900 nukleotidov) a vo veľkej podjednotke 5S rRNA (120 nukleotidov), 5,8S rRNA (160 nukleotidov) a 28S rRNA (4 700 nukleotidov). 16S rRNA je zložkou A miesta ribozómu.
Proteíny
Ribozómy sa skladajú tiež z proteínov. Prokaryotické ribozómy obsahujú 55 proteínov (21 v malej a 34 vo veľkej podjednotke), eukaryotické majú dokonca 82 proteínov (33 v malej a 49 vo veľkej podjednotke). Bielkoviny ribozómov majú prevažne kladný náboj, ale nie sú blízke histónom. Obsahujú veľký počet lyzínových a arginínových zvyškov. Mnohé z nich majú dosť neobvyklú štruktúru s dlhými roztiahnutými ramenami obohatenými o bázické bielkoviny. Tie pravdepodobne umožňujú zvinutie negatívne nabitej fosfátovej kostry rRNA do kompaktnej priestorovej štruktúry.
Ribozomálne proteíny možno rozdeliť na
- jednotkové – vyskytujú sa v každom ribozóme v jednej až dvoch kópiách. Majú hlavne štruktúrnu funkciu – tvoria kostru obidvoch podjednotiek.
- okrajové – nevyskytujú sa v každom ribozóme, frekvencia ich výskytu je 0,5 až 0,8 molekuly na jeden ribozóm
- zlomkové – frekvencia ich výskytu je menšia ako 0,5
Niektoré ribozomálne proteíny sú asociované so špecifickými väzobnými miestami:
- A miesto – jeho súčasťou sú proteíny L1, L5, L7, L12, L30, L33
- P miesto – jeho súčasťou sú proteíny L7, L12, L14, L18, L24, L33
Polyzómy
U prokaryotov aj eukaryotov sa na jednej mRNA obvykle nachádza niekoľko ribozómov, čím sa syntéza proteínov značne urýchľuje. Hneď, ako sa ribozóm posunie po mRNA natoľko, že celkom uvoľní väzobné miesto, na toto miesto sa naviaže ďalší ribozóm. Jednotlivé ribozómy sú na mRNA vzdialené od seba približne 80 báz. Ribozómy pokračujú v syntéze proteínu až kým mRNA nie je degradovaná, čo sa stáva po niekoľkých minútach (prokaryotická mRNA) až po viac ako 10 hodinách (eukaryotické mRNA pre proteíny potrebné vo veľkých množstvách).
Referencie
- ROZSYPAL, Stanislav. Úvod do molekulární biologie. Zväzok 1. Brno : [s.n.], 1998. 3 zv. (česky)
- MIŠÚROVÁ, Eva; SOLÁR, Peter. Molekulová biológia. [s.l.] : Univerzita Pavla Jozefa Šafárika v Košiciach, 2007. ISBN 978-80-7097-671-5.
- ŠKÁRKA, Bohumil; FERENČÍK, Miroslav. Biochémia. Bratislava : Alfa/Vydavateľstvo technickej a ekonomickej literatúry, 1983. Kapitola 8.2. Ribozómy, s. 225.
- ALBERTS, Bruce; BRAY, Dennis; JOHNSON, Alexander, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter Základy buněčné biologie. Redakcia Miranda Brownová, Eleanor Lawrenceová, Valerie Nealová, Anne Vinnicombeová; preklad Arnošt Kotyk, Bohumil Bouzek, Pavel Hozák; ilustrácie Nigel Orme. 2. vyd. Ústí nad Labem : Espero Publishing, © 1998. ISBN 80-902906-2-0. Kapitola Od DNA k proteinu, s. 223 – 232. (česky)
- BRESLER, S. J.. Molekulární biologie. Preklad Stanislav Zadražil. Praha : ACADEMIA, 1979. (česky)
- Bruce Alberts et al (2004). Essential Cell Biology. Garland Science.
- Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell, 4rd edition [online]. Dostupné online.
- MANGIAROTTI G., SCHESSINGER D.: J. mol. Biol. 20, 123 (1965)
- KAEMPFERVR., MESELSON M.: J. Mol. Biol. 31,277 (1968)

Autor alamo So jan 28th 2017 at 9:44 pm
aký vlastne majú pôvod?
bunky si ich vytvorili sami, alebo sú to ešte viac „zdomestifikovaný“ simbionti, na spôsob mitochondrií?
sú o tom nejaké teórie?
keď som to naposledy skúšal googliť, beznádejne som sa stratil v odborných termínoch a google preklade..
Autor Adhara Ne jan 29th 2017 at 1:48 pm
Neviem. No pokiaľ mi je známe, o endosymbióze sa v prípade ribozómov nikdy neuvažovalo. Veď jednak nemajú membrány a jednak život bez ribozómov, teda pred ribozómami, by bol ťažko funkčný. Môj osobný názor je, že ribozómy aspoň v nejakej zjednodušenej forme museli predchádzať bunkám.
Autor alamo Ne jan 29th 2017 at 9:18 pm
už som si spomenul, odkiaľ sa to na mňa „nalepilo“
https://en.m.wikipedia.org/wiki/Ribosome#Origin
lebo „kosmo“, s času na čas nás tam chytne mánia debatovať o mimozemskom živote, možnostiach a nemožnostiach jeho vzniku atď
pred časom ma tak zaujali články o „RNA svete“
ribozómy by z neho boli z jedným z mnohých pozostatkov, a fungovali kedysi ako samostatný „replikátor“, ktorý sa naučil spolupracovať s inými spolu, dávno pred vznikom DNA
Autor alamo Št feb 2nd 2017 at 11:35 am
vždy keď sme o možnostiach vzniku života na kosmo.cz debatovali, točilo sa to okolo otázky „ako vznikol prvotný replikátor?“
„jeden stroj – organická makromelekula schopný postaviť svoju kópiu“
čo ak je to príliš zjednodušujúca laická predstava?
náhodné samozloženie niečho takého, je naozaj krajne nepravdepodobné
ako som sa v tom, po prečítaní tohto tvojho článku prehraboval, napadlo ma že to bolo asi pekelne zložitejšie
čo ak musel vzniknúť nie jeden, ale celý „komplex“ na vzájom zdanlivo nezávislých jednotiek, samostatne ale omnoho oveľa jednoduchších než „jediný dokonalý“, s ktorých každá tvorila svoju časť prvotného metabolizmu?
„replikátor jednotka a“ metabolizuje vstupný materiál, vzniká „odpadný produkt“, ten metabolizuje zase „jednotka b“ za vzniku iného „produktu“, ktorý zase používa „jednotka c“ ktorej výstup potrebuje na začiatku „jednotka a“
pravdepodobne, v takomto cykle ich boli zapojené možno aj aj desiatky, a väzby medzi nimi (vzájomné výmeny) neboli iba „kruhové“ ale aj „priečne“, mohli by sa pridávať nové „kroky“, iné sa vylučovať
mohlo by takéto niečo, teoreticky fungovať rozptýlené v „prapolievke“ na veľké vzdialenosti od seba, bez toho aby to muselo byť oddelené od okolitého prostredia membránou a tvoriť bunku?
Autor Adhara Pi feb 3rd 2017 at 3:39 pm
Takto komplikovane ako popisuješ to funguje aj v súčasnosti. Nijaká molekula sa nevie replikovať sama, bez pomoci iných. Vždy je to o spolupráci a myslím, že aj vždy bolo. Práveže predstava jednej molekuly vytvárajúcej kópiu samej seba mi pripadá ako veľké zjednodušenie, ku ktorému by sa možno dalo dospieť evolúciou.
V prednáške „Zázrak, že sme tu“ túto problematiku trochu rozoberám. Isteže sa nedá očakávať, že v prvotnej polievke oceánov mladej Zeme by vznikla súčasná E. coli ani súčasný Methanococcus. Isteže tomu predchádzalo niečo omnoho jednoduchšie, neefektívnejšie „položivé“. Aj vo vzniku niečoho omnoho jednoduchšieho však vidím stále obrovskú nepravdepodobnostnú bariéru. Tak napríklad: náhodný vznik jednej výkonnejšej molekuly mi pripadá pravdepodobnejší ako náhodný vznik hromady menej výkonných, ktoré spoločne robia to isté, čo jedna výkonnejšia. Polymerizácia bez katalyzátora je totiž rovnako málo pravdepodobná pri zmysluplných ako aj pri nezmysluplných sekvenciách. Ak neboli žiadne katalyzátory, čo aj anorganické, potom platí „čím menej zapojených aminokyselín, tým väčšia šamca, že také niečo samovoľne vznikne“. No lenže čím menej aminokyselín (alebo báz), tým väčší užitočnejšie musia byť tie, čo zapojené sú… Nevraviac o tom, že tie menej výkonné sa ešte musia spolu ocitnúť na jednom mieste, čo je tiež riadna náhodička.
Autor Tomi Lee TRNíK So feb 4th 2017 at 3:04 pm
Len dodám, že k vzniku života treba HLAVNE pridať miliardu rokov. Všetci čo tvrdia, že si nevedia PREDSTAVIŤ, že život mohol vzniknúť len tak, by si mali skúsiť predstaviť čo bude za 100 či 1000 rokov. _ _ _
To si podla mňa nikto nevie ani len predstaviť, takže miliarda rokov je dostatočný čas, aby doslova aj z ničoho vzniklo čokoľvek. Mňa napríklad už vznik života vôbec neprekvapuje, po tom čo som túto oblasť trocha preskúmal pri písaní o Zmysle Života._ _ _
Inak povedané, aj dnes niektoré (všetky) živočíchy svojim telom a správaním sú AŽ neuveriteľne prispôsobené svojmu prostrediu, ale táto komplikovanosť, nevznikla cez noc, ale trvalo to milióny rokov. A tak aj komplikovanosť samotnej bunky, je dôsledkom času, ktorý všetko menej funkčné vyradil z obehu.
Autor Adhara So feb 4th 2017 at 6:10 pm
Ehm, vznik života ale netrval celú miliardu rokov. Len tristo miliónov. :-) Ale OK, viem, že to vyzerá ako chytanie za slovíčka. Idem sa teda zamerať na podstatu, že na to treba naozaj VEĽA času.
Čas je pri vzniku života dvojsečná zbraň. Jedna vec je, že aj niečo málo pravdepodobné sa stáva pravdepodobnejším, keď tomu dáme viac času (preto som niektoré svoje projekty ani po viac ako desaťročí neúspešných pokusov nevzdala). Druhá vec ale je, že bez ochrany membrán, v silne rádioaktívnom prostredí mladej Zeme, sú polyméry strašne nestabilné. To znamená, že keď nám vznikne jeden aspoň trochu funkčný polymér, nemôže si dovoliť čakať 20 000 rokov, kým sa mu niekde poblíž sformuje „parťák“, aby mohli spoločne zostavovať metabolizmus.
Autor Tomi Lee TRNíK Po feb 6th 2017 at 5:11 pm
malá VEĽKÁ otázka: máš záujem o moju odpoveď, kde budem vysvetľovať ako mohol podla mňa vzniknúť život?
Autor alamo Po feb 6th 2017 at 9:52 pm
neviem ako to presne „podať“..
aký je vlastne vzťah medzi „metabolizmom“ a „replikáciou“?
replikáciu zvládajú predsa aj neživé veci kryštály, kvázi kryštály ílov
http://www.osel.cz/543-jak-je-to-s-temi-zivymi-jily.html
aj „mydlo“ tvorí lipidové membrány spontánne, a vďaka tomu vlastne funguje
mám dojem ako keby „čistá“ replikácia niečo, čomu ako zdroj pohybu „pohonu“ dostačuje Brownov pohyb.. žiadny metabolizmus na to, akoby ani nebolo treba, aspoň teraz mám taký dojem
metabolizmus hmm.. čo všetko sa pod to zahrňuje?
ako laik tam identifikujem, hlavne dve rozdielne veci
a, energetický príjem a výdaj (napríklad pre aktívny pohyb)
b, schopnosť budovať si vlastné stavebné prvky
..
existuje čosi ako teória „malej panspermie“
podľa nej počas veľkého bombardovania, došlo k nejakému prenosu niečoho (nie „hotového života“, iba nejakej jeho „súčiastky“) medzi mesiacmi veľkých planét s podľadovými oceánmi, ktoré ale majú podľa niektorých modelov dlhodobo stabilnejšie podmienky než Zem
čo malo spôsobiť čosi ako“zamiešanie“ a následne „revolúciu“ v omnoho bohatšom a pestrajšom prostredí
bohužiaľ neviem teraz nájsť zdroj s ktorého to mám