


Slovník genetických hesiel, ktoré som spracovala pre Wikipédiu. Môžu byť užitočné tak záujemcom o prírodné vedy, ako aj čitateľom knižnej série Nula kelvinov. Vybrala som také pojmy, ktoré sú aspoň okrajovo spomenuté v deji jednej či druhej z kníh. Uvedomujem si však, že neposkytujú komplexné základy v téme expresie génov, ba ani všetkých pojmov spomínaných v knihách – sú totiž limitované výberom z toho, čo som pre Wikipédiu spracovala ja osobne.
Heslá sú radené abecedne a možno ich čítať samostatne. Kurzíva značí, že heslo je rozpísané samostatne (nahrádza wikilinku).
Expresia génu
(spoluautori hesla Saskardin a IP 88.212.16.234)
Gén chápaný v zmysle úseku DNA prepisovaného do proteínov môže mať všetky sekvencie funkčné a predsa sa nemusí prejavovať. V organizmoch nedochádza k prejavom všetkých génov súčasne. U mnohobunkových organizmov dochádza v rôznych orgánoch k prejavom rôznych génov, hoci všetky bunky majú takmer identickú DNA a všetky teda nesú rovnaké gény. Prejavovanie génu sa nazýva expresia génu. Je ovplyvňovaná rozličnými faktormi. Expresia génu sa môže v závislosti od okolností alebo od štádia ontogenézy zapínať a vypínať.
Niektoré gény sa prejavujú vo všetkých bunkách daného organizmu. Ide o gény riadiace chod každej bunky, napríklad ribozomálne gény. Tieto gény sa nazývajú „housekeeping gény“. Iné gény sú špecifické len pre určité tkanivo alebo pletivo, napríklad gén pre hemoglobín v krvných bunkách.
Genetický kód
Ako informácia pre zaradenie určitej aminokyseliny do vznikajúceho peptidového reťazca slúžia v DNA (a takisto v jej transkripte, RNA) tri vedľa seba ležiace nukleotidy. Táto trojica nukleotidov – tzv. triplet sa nazýva kodón. Keďže bázy sú štyri a každá z nich sa môže nachádzať na jednej z troch pozícií, malo by existovať 64 rôznych kodónov. Z nich však tri nekódujú nijakú aminokyselinu a ich prítomnosť v RNA naznačuje, že v tomto mieste je potrebné syntézu proteínu ukončiť. Aj tak však zostáva 61 rôznych kodónov, pričom poznáme len 21 aminokyselín, ktoré organizmy používajú pre proteosyntézu. Rozličné kodóny však môžu kódovať tie isté aminokyseliny. Len metionín, tryptofán a selenocysteín majú svoje kodóny jedinečné. Ostatné aminokyseliny sú kódované dvoma, štyrmi až šiestimi rozličnými kodónmi. Kvôli tejto vlastnosti sa genetický kód nazýva degenerovaný.
Gén
(spoluautori hesla Saskardin, IP 88.212.16.234)
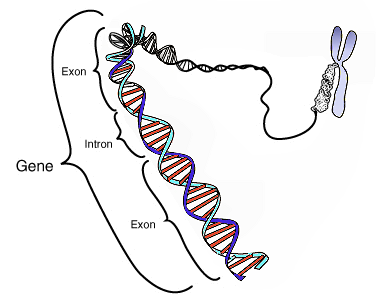
Umiestnenie génu v chromozóme a jeho časti intrón a exón. Zdroj obrázku
Gén je úsek (sekvencia) DNA alebo RNA, ktorá kóduje informáciu na tvorbu nejakého produktu. V užšom zmysle gén kóduje tvorbu jednej bielkoviny. Existuje množstvo rôznych definícií génu, často upravených na potreby jednotlivých vedných disciplín. Gén je základná funkčná jednotka dedičnosti.
Gén, ktorý kóduje proteín, je zložený z tripletov – kodónov, kde každý triplet predstavuje informáciu o zaradení jednej aminokyseliny. Zároveň musí gén obsahovať aj sekvencie umožňujúce jeho reguláciu. Patria sem sekvencie označujúce miesto začiatku a konca transkripcie génu. Tieto kodóny nekódujú aminokyseliny, ale poskytujú informácie, bez ktorých by nebol možný úspešný priebeh proteosyntézy. Úseky kodónov bez funkčných regulačných sekvencií sa nazývajú pseudogény.
Gény tvoria veľkú časť DNA baktérií, no len malú časť v DNA eukaryotov. Eukaryotická DNA totiž obsahuje rozsiahle nekódujúce sekvencie medzi génmi, ale aj vnútri nich. Nekódujúca sekvencia vnútri génu sa nazýva intrón. Aby z génu vznikol správny produkt, musí byť intrón z pre-mRNA vystrihnutý a zostávajúce kódujúce časti RNA spojené. Kódujúca časť génu sa nazýva exón. Časť DNA tvorená exónmi sa nazýva kódujúca DNA.
Gén v širšom zmysle
Génom možno nazvať hocijaký úsek DNA, ktorý je prepisovateľný do podoby RNA, ktorá následne plní v bunke nejakú funkciu. Tou funkciou nemusí byť len syntéza bielkovín. Niektoré druhy RNA dokážu vykonávať funkciu už samé osebe, hoci predtým neraz ešte musia podstúpiť úpravy. Takáto RNA sa nazýva funkčná RNA a matrica na jej vytvorenie je gén v širšom zmysle.
Kodón
(Spoluautori hesla IP 195.28.68.82, AtonX, Ošo, Bronto)
Kodón je sekvencia troch nukleotidov (nazývaných v tejto súvislosti triplet) v molekule DNA alebo mRNA, ktoré určujú zaradenie konkrétnej aminokyseliny do polypeptidového reťazca. Určitý kodón kóduje len jednu aminokyselinu, avšak určitá aminokyselina môže byť kódovaná viacerými, rôznymi kodónmi, napríklad arginín, serín a leucín sú kódované každá až šesticou rôznych kodónov. Kodóny kódujúce tú istú aminokyselinu sa nazývajú synonymné; mutácia, ktorá zmení kodón na iný synonymný kodón, sa nazýva tichá mutácia.
Kodóny sa v DNA vyskytujú na tzv. pozitívnom z dvoch komplementárnych reťazcov génu, a na mRNA. Sekvencia mRNA sa však odlišuje od pozitívneho reťazca génu tým, že namiesto tymínu sa v nej nachádza báza uracil. Funkcia kodónu spočíva v tom, že v procese translácie sa naň pripojí antikodón, čo je sekvencia troch komplementárnych báz v molekule tRNA. Po tomto spárovaní tRNA, ktorá daný antikodón nesie, pripojí svoju aminokyselinu k tvoriacemu sa peptidovému reťazcu.
Väčšina kodónov je čítaná v rôznych organizmoch od baktérií až po človeka rovnako. Hovoríme preto o tzv. univerzálnom genetickom kóde. Existujú však aj takzvané minoritné kodóny, ktoré sa vyskytujú zriedkavo a sú špecifické pre niektoré skupiny organizmov alebo pre niektoré bunkové organely.
Špeciálny typ kodónu:
• Iniciačný (AUG) – označuje začiatok pre syntézu bielkovín pri translácii a kóduje bielkovinu metionín
• Terminačný (UGA, UAA, UAG) – označuje koniec syntézy bielkovín pri translácii a nekóduje žiadnu bielkovinu. UAG však môže byť niekedy čítaný ako kodón pre selenocysteín.
Mitochondriálna DNA
Mitochondriálna DNA alebo mitochondriová DNA (skratka mtDNA) je genetická informácia, ktorá sa nachádza v mitochondriách. Podieľa sa na mimojadrovej dedičnosti. Z celkového množstva DNA v eukaryotickej bunke pripadá na mitochondriálnu DNA obvykle 1 % – 0,5 %.
Mitochondriálna DNA je tvorená 2 až 6 molekulami DNA, ktoré sú obvykle dvojreťazcové a uzavreté do kruhu, podobne ako chromozómy prokaryotov. Na základe toho sa usudzuje, že mitochondrie boli kedysi samostatnými prokaryotickými organizmami, ktoré sa stali endosymbiontami eukaryotických buniek. Genóm mitochondrií prechádzal evolúciou, počas ktorej sa z neho niektoré gény stratili či premiestnili do jadra. Molekulová hmotnosť mtDNA sa pohybuje rádovo v 10 000 000 Da.
Ľudská mtDNA sa skladá z 16 569 deoxyribonukleotidov a obsahuje 37 génov. Medzi nimi (tak ako medzi génmi mtDNA iných organizmov) sú aj dôležité gény kódujúce podjednotky dýchacích enzýmov: citochrómy a oxidázy. Mitochondriálna DNA je teda nevyhnutná pre život väčšiny eukaryotických buniek. Pre úspešnú syntézu bielkovín kódovaných v mtDNA zase musí byť do mitochondrie transportovaných množstvo produktov jadrových génov.
Kód mtDNA sa v mnohom odlišuje od jadrovej DNA. Jej gény sú na rozdiel od jadrovej usporiadané veľmi úsporne a dôležitý je v nich každý bázový pár. Posledný nukleotid jedného génu môže dokonca v niektorých prípadoch byť zároveň prvým nukleotidom ďalšieho génu. Ďalšou zvláštnosťou je sekvencia UGA, ktorá v jadrovej RNA znamená koniec syntézy peptidového reťazca (stop kodón), ale v mitochondrii táto sekvencia kóduje aminokyselinu tryptofán. Transkripciu mitochondriálnych génov zabezpečuje zvláštna mitochondriová RNA-polymeráza.
Mitochondriálna DNA sa na rozdiel od jadrovej, v ktorej sa polovica zdedí po otcovi a polovice po matke, dedí vo väčšine prípadov iba po matke. Takáto dedičnosť sa nazýva materálna.
Mutácia
Mutácia je zmena genotypu jedinca. Môže prebehnúť spontánne, tj. bez vonkajšej príčiny, alebo pod vplyvom pôsobenia prostredia (indukovaná mutácia). Pokiaľ sa táto zmena nevráti do pôvodného stavu (procesom reverzie alebo reparácie mutácií), prenáša sa na všetky bunky (resp. bunkové organely, pokiaľ zmena nastala v mimojadrovej DNA), ktoré vzniknú delením pôvodnej zmutovanej bunky.
Mutácie sa rozlišujú z viacerých hľadísk. Podľa rozsahu jadrových štruktúr, ktoré postihujú, ich možno rozdeliť na genómové, chromozómové a génové. Mutácia môže postihnúť telovú bunku (somatickú), alebo pohlavnú (gamétu). V druhom prípade sa mutácia stáva dedičnou a prenáša sa na ďalšie generácie organizmov. Dedičné mutácie sú jedným z hlavných procesov dedičnej premenlivosti a dôležitým faktorom evolúcie. Na druhej strane, somatické mutácie postihujú len daný organizmus a to v o to väčšom rozsahu, v o čo skoršej fáze jeho vývinu mutácia nastala. Somatické mutácie môžu byť napríklad podkladom rakovinových ochorení, alebo môžu zmeniť vzhľad (fenotyp) jedinca.
Pokiaľ majú zmutované jedince odlišný výzor ako nezmutované, hovoríme o morflologických mutáciách. Ak mutácia nie je zlúčiteľná so životom (jedinec odumrie už v štádiu zygoty alebo v skorej embryonálnej fáze) či spôsobí úmrtie organizmu, nazýva sa letálna mutácia. Rôzne veľké mutácie majú rôzny postih na skupiny organizmov. U cicavcov je napríklad znásobenie chromozómovej sady (polyploidia) nezlúčiteľná so životom, ale mnohé kultúrne rastliny majú svoje chromozómové sady v polyploidnom stave. V niektorých prípadoch sa však naopak mutácia nemusí na fenotype vôbec prejaviť a nemá ani vplyv na prežívanie jedincov.
Primer
Primer (čítaj prajmer) alebo novšie primér je úsek ribonukleovej kyseliny (RNA), ktorý zohráva dôležitú úlohu v procese replikácie DNA. Je to krátky reťazec pozostávajúci len z niekoľkých ribonukleotidov – oligonukleotid, na ktorého 3′-OH skupinu dokážu replikačné enzýmy zo skupiny DNA-polymeráz pridávať deoxyribonukleotidy a zahájiť tým tvorbu nového dcérskeho reťazca DNA. DNA-polymerázy totiž dokážu pridávať deoxyribonukeotidy len k už existujúcemu reťazcu nukleovej kyseliny. Preto je primer dôležitý na začiatku procesu vlastnej replikácie DNA.
Primer vzniká pôsobením enzýmu primázy (RNA-primázy). U eukaryotov má primázovú aktivitu (je schopná tvoriť primery) DNA polymeráza α. Na matrici DNA, ktorá má smer 3′-5′ postačí na replikáciu celého vlákna len jeden primer čím vzniká tzv. vedúci reťazec (leading strand). Avšak na vlákne so smerom 3′-5′ sa nový reťazec DNA syntetizuje v podobe krátkych (1000-2000 báz) úsekov, tzv. Okazakiho fragmentov, z ktorých syntéza každého začína tvorbou nového primeru. Pri replikácii tohto tzv . zoastávajúceho reťazca (lagging strand) sa teda musí vytvoriť množstvo primerov. Po pripojení niekoľkých deoxyribonukleotidov už primer nemá nijakú funkciu a tak sa z oboch reťazcov odstráni pôsobením enzýmu ribonukleázy H.
V širšom zmysle je primer akýkoľvek voľný 3′-OH koniec nukleovej kyseliny (teda aj DNA), na ktorý sa môžu naviazať DNA polymerázy.
Restrikčná endonukleáza
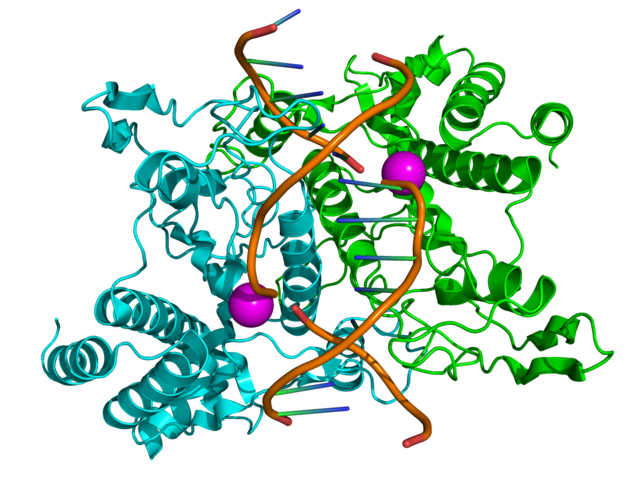
EcoRI, jedna z najznámejších restrikčných endonukleáz pochádzajúca z baktérie Escherichia coli, pri štiepení DNA. Zdroj obrázku
Restrikčná endonukleáza (iné označenia: restrikčná nukleáza, restrikčný enzým, restriktáza) je enzým, ktorý štiepi DNA v miestach rozpoznaných vďaka krátkemu úseku jej sekvencie. Každý druh restrikčnej endonukleázy má svoju špecifickú rozpoznávaciu sekvenciu, ktorá sa líši od iných druhov restriktáz.
Sekvencia rozpoznávaná restrikčnými endonukleázami má dĺžku obvykle 4-8 bázových párov. Niektoré restriktázy rozštiepia v miestach ležiacich presne oproti sebe, iné rozštiepia druhé vlákno na pozícii posunutej o niekoľko nukleotidov voči prvému, takže na konci takto rozštiepeného dvojvlákna vznikne krátky jednovláknový úsek. Restrikčné endonukleázy sú veľmi používané v biologických výskumoch a pri technikách využívajúcich rekombinantné DNA. Väčšina restriktáz používaných na tento účel bola izolovaná z baktérií.
RNA-polymeráza
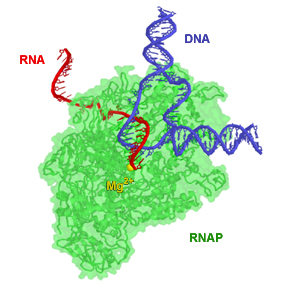
RNA-polymeráza počas predlžovania reťazca RNA. RNA-polymeráza na krátkom úseku štiepi vodíkové väzby medzi bázami DNA, aby oddelila od seba jej vlákna a mohla použiť jedno z nich ako matricu pre syntézu RNA. Zdroj obrázku
(Spoluautor hesla Saskardin)
RNA-polymeráza je enzým, ktorý katalyzuje vznik fosfodiesterovej väzby medzi ribonukleotidmi a tým vytvára ribonukleovú kyselinu (RNA). Je to hlavný enzým podieľajúci sa na procese transkripcie (prepisu) genetickej informácie. Produktom jej činnosti je reťazec RNA označovaný ako primárny transkript.
RNA-polymerázy sú podobné DNA-polymerázam v tom, že vyžadujú vlákno DNA (vzácne RNA) ako matricu, na ktoré pripájajú nukleotidy na základe princípu komplementarity. Tiež dokážu syntetizovať reťazec RNA len v smere 5′-3′. Na rozdiel od DNA-polymeráz ale RNA polymeráza nekontroluje pred pripojením ďalšieho nukleotidu správnosť zapojenia predchádzajúceho nukleotidu. Výhodou tejto jej vlastnosti je, že nepotrebuje primer. Nevýhodou je, že takýto prepis génu je menej presný, než by bolo komplementárne vlákno DNA.
U baktérií a archeí je známych len po jednom type RNA-polymerázy. Eukaryoty majú tri rôzne RNA-polymerázy označené rímskymi číslicami ako I, II a III.
Telomeráza
Telomeráza je enzým, ktorý slúži na replikáciu telomér, koncov eukaryotických chromozómov. Replikačný mechanizmus, ktorý zreplikuje takmer celý chromozóm, totiž nie je schopný zreplikovať jeho koniec. Problém nastáva v tzv. „oneskorujúcom sa vlákne“ (lagging strand), pre ktoré sa na konci chromozómu už nemôže vytvoriť primer. Na účely jeho doreplikovania preto existuje osobitný enzým telomeráza, ktorý má vlastnosti reverznej transkriptázy. To znamená, že vytvára DNA podľa matrice vo forme RNA. Táto RNA matrica je štruktúrnou súčasťou samotnej telomerázy.
Bez telomerázy by sa konce eukaryotických chromozómov pri každej replikácii DNA skracovali. Po určitom počte delení, ktorý nazývame Hayflickov limit, sa preto bunky bez aktivity telomerázy už nemôžu ďalej deliť. Telomeráza preto pracuje v bunkách, ktoré potrebujú väčší počet delení než dovoľuje Hayflickov limit, čo sú napríklad embryonálne bunky.
Umelá stimulácia tvorby telomerázy by mohla byť jedným z faktorov, ktoré by pomohli oddialiť starnutie organizmu. Na druhej strane, až 90 % nádorov sa vyznačuje telomerázovou aktivitou, vďaka čomu majú nádorové bunky prakticky neobmedzený počet delení. Inhibícia telomerázy by preto naopak mohla pomôcť pri liečbe rakoviny.
Niektoré organizmy môžu mať viac bunkových delení, ako dovoľuje aj v neprítomnosti telomerázy. Takéto organizmy využívajú alternatívne riešenia replikácie koncov chromozómov. Kvasinky Schizosaccharomyces pombe napríklad svoje chormozómy cirkularizujú, čím sa stávajú podobnými bakteriálnym chromozómom, ktoré telomerázu nepotrebujú.
Časť telomerázy je u ľudí kódovaná génom s označením TEP1. Nachádza sa na chromozóme 14. Gén sa exprimuje v niektorých tkanivách vyvíjajúceho sa embrya, v neskoršom vývoji v zdravých tkanivách tvorba enzýmu celkom ustane.
Zdroje
- ROSYPAL, Stanislav. Úvod do molekulární biologie. tretie. vyd. Brno : [s.n.], 1998. (česky)
- Nukleové kyseliny [online]. Biopedia.sk, [cit. 2011-01-13]. Dostupné online.
- http://is.muni.cz/th/151360/prif_b/bc_MB.txt
- DNA deoxyribonukleová kyselina [online]. Encyklopédia Vševěd, [cit. 2011-01-13]. Dostupné online. (česky)
- ALBERTS, Bruce; BRAY, Dennis; JOHNSON, Alexander, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter Základy buněčné biologie. Redakcia Miranda Brownová, Eleanor Lawrenceová, Valerie Nealová, Anne Vinnicombeová; preklad Arnošt Kotyk, Bohumil Bouzek, Pavel Hozák; ilustrácie Nigel Orme. 2.. vyd. Ústí nad Labem : Espero Publishing, © 1998. ISBN 80-902906-2-0. (česky)
- Nukleové kyseliny a proteosyntéza [online]. [Cit. 2011-02-09]. Dostupné online.
- THOMPSON, James S.; THOMPSONOVÁ, Margaret W.. Klinická genetika. Preklad Viliam Izakovič. štvrté. vyd. Martin : Osveta, 1988. 440 s.
- MIŠÚROVÁ, Eva; SOLÁR, Peter. Molekulová biológia. [s.l.] : Univerzita Pavla Jozefa Śafárika v Košiciach, 2007. ISBN 978-80-7097-671-5.

Napísať odpoveď